Самое время наполнить корзину
Как ящерица отбрасывает хвост (автотомия)
 Впервые приобретая ящерицу, новый владелец озадачен многими вопросами и озабочен многими проблемами. Как создать ей все необходимые условия для содержания? Как обеспечить комфорт своему новому питомцу? Как понять, что со здоровьем эублефара все в порядке? А одним из главных вопросов является следующий – почему ящерица может потерять хвост, и как это предотвратить? Ответ на него довольно прост – будьте предельно аккуратны в обращении с питомцем, не дергайте и не тяните его за хвост, а также, постарайтесь свести к минимуму стрессовые ситуации.
Впервые приобретая ящерицу, новый владелец озадачен многими вопросами и озабочен многими проблемами. Как создать ей все необходимые условия для содержания? Как обеспечить комфорт своему новому питомцу? Как понять, что со здоровьем эублефара все в порядке? А одним из главных вопросов является следующий – почему ящерица может потерять хвост, и как это предотвратить? Ответ на него довольно прост – будьте предельно аккуратны в обращении с питомцем, не дергайте и не тяните его за хвост, а также, постарайтесь свести к минимуму стрессовые ситуации.
А пока вы готовитесь к покупке нового члена семьи или уже наслаждаетесь его обществом, прочитайте о том, как происходит автотомия - потеря хвоста у ящериц – и его регенерация.
«Автотомия (частичная или полная потеря конечности по желанию владельца) – это ответ на механические или визуальные стимуляции, имеющие отношения к таким факторам, как межвидовая конкуренция или агрессия, предпринимаемая попытка атаковать другое существо или абиотические влияния по типу экстремальных изменений температуры. Автотомия очень эффективна как стратегия, ибо она очень распространена среди различных групп позвоночных и беспозвоночных – у ящериц, морских звёзд, пауков, амфибий и млекопитающих. В случае многих животных, отброшенная конечность не активна настолько, чтобы это можно было заметить. Но в случае с ящерицами, все происходит с точностью до наоборот.
Автотомия у ящериц эффективна в случае нападения хищника потому, что отброшенный хвост приковывает к себе внимание, и хищник фокусируется на нем и на том, чтобы съесть его, а не гнаться за хозяином хвоста.
Экологическим и эволюционным аспектам отбрасывания хвоста у ящериц было уделено много внимания в различных исследованиях. Большинство из них фокусировалось на последствиях потери хвоста для ящерицы (поведение, биомеханика, энергетика и т.д.) Например, скорость бега может значительно увеличиться после автотомии, что повышает шансы ящерицы убежать от преследователя. Однако потеря хвоста плохо сказывается на способности прыгать, совершать манёвры в воздухе и сохранять стабильность при ползании по вертикальным поверхностям. Это также может негативно повлиять на социальные качества и приспособленность к жизни в общем (репродуктивность, потенциал к выживанию, энергичность и общее поведение). Но выгода от избавления от хищника, очевидно, превышает отрицательные последствия потери хвоста, а недавние исследования вообще ставят под вопрос уровень того, насколько дорого потеря хвоста обходится его обладателям.
Сам по себе хвост также может продемонстрировать интересные поведенческие паттерны, как только отделится от тела. Например, последняя работа указала на способность хвостов леопардовых гекконов подпрыгивать и переворачиваться в дополнение к возможности двигаться из стороны в сторону. Тем не менее, немного известно о механизмах контроля, лежащих в основе таких движений.
Используя электромиографию, ученые исследовали модели движения хвостов эублефаров, зависящие от времени, на живых организмах и в 4 типах среды (двух отдаленных и двух ближайших) сразу после автотомии.
Они тестировали гипотезу о том, что расхождение в движениях является исключительно результатом перекликающихся моделей внутри хвоста. Было обнаружено, что продолжительность взрыва, но не продолжительность цикла, ритмических сокращений достигла плато (точки спокойствия) после автотомии на отметке примерно 150. Скорее всего, это происходит в связи с физиологическими изменениями, имеющими отношение к мышечной усталости и ишемии (уменьшение кровоснабжения участка тела, органа или ткани вследствие ослабления или прекращения притока артериальной крови). В том, что касается переворотов и прыжков, продолжительность взрыва и цикла не обнаружила регулярной закономерности.
Коэффициент вариации в моделях движения был значительно больше для прыжков и переворотов, чем для ритмичных покачиваний. Это подтверждает вывод о том, что различные поведения хвоста происходят не в связи с перекликающимися источниками энергии для совершения движений, но полностью зависят от независимых нейронных цепей. Сигнал, контролирующий прыжки и перевороты, может быть модифицирован сенсорной информацией из окружающей среды.
Наконец, ученые обнаружили, что прыжки и перевороты вызваны использованием относительно синхронной активности между двумя сторонами хвоста. Попеременная активация левой и правой частей хвоста, напротив, выражается в ритмических покачиваниях. Механизм, лежащий в основе такого изменения поведения хвоста, можно сравнить с изменениями в движениях опорно-двигательного аппарат у позвоночных.
Каковы же истоки сигналов, заставляющих хвост двигаться подобным образом? Результаты исследования показали, что как минимум две независимые нейронные цепи активны в хвосте. Одна обеспечивает источник регулярно повторяющихся ритмичных сигналов, присутствующих во время движений хвоста после автотомии. Другая – кратковременна и отвечает за сложные прыжки и перевороты. Главным вопросом, оставшимся без ответа, является следующий – каким образом активируется дополнительная нейронная цепь? Есть три возможных объяснения: 1) это либо случайный запуск движения до тех пор, пока не наступит усталость; 2) либо чувствительность прохода (места, где хвост оторвался) к условиям окружающей сред, либо 3) вытекающая из предыдущего утверждения чувствительность прохода к проприорецепторам мышц (мышечные веретена и сухожильные органы гольджи)».
Не правда ли, удивительно? Ваша домашняя ящерица стала еще любопытнее.
А что насчет регенерации? Как проходит процесс восстановления тканей и, как следствие, всей части тела?
«Эпиморфная регенерация заключается в восстановлении потерянных тканей и структур посредством агрегации размножающихся клеток, известных, как бластема. Самый выдающимся примером эпиморфной регенерации среди амниотов являются ящерицы, способные восстанавливать хвост.
Эпиморфоз – способ регенерации от раневой поверхности, который заключается в отрастании нового органа от ампутированной поверхности. Эпиморфная регенерация может быть типичной, если восстановившийся после ампутации орган не отличается от неповрежденного, или атипичной, когда восстановившийся орган по форме или структуре отличается от нормального. Примером типичной регенерации служит восстановлении конечности у аксолотля (личинки амбистомы) после ампутации.
Несмотря на то, что восстановление хвоста часто изучается в контексте экологических затрат и выгод, деталей, известных о процессах, происходящих в тканях, недостаточно. Был проведен опыт с целью исследования анатомических и гистологических процессов, которые сопутствуют восстановлению хвоста у леопардовых гекконов.
Структура хвоста и состав тканей были изучены различными путями сразу же после потери хвоста и обнаружили устойчивую модель регенерации. Удаление хвоста ведет за собой постоянную цепочку морфологических и гистологических изменений. (Гистология — раздел биологии, изучающий строение, жизнедеятельность и развитие тканей живых организмов. Обычно это делается рассечением тканей на тонкие слои и с помощью микротома). Сразу после потери хвоста идет период заживления раны без каких-либо видимых признаков регенеративных наростов. В течение этого латентного (скрытого, внешне не проявляющегося) периода базальные клетки эпидермиса делятся и постепенно покрывают рану (Базальный слой — слой клеток в основании эпидермиса, прилегающий к дерме. Клетки базального слоя, размножаясь, обеспечивают постепенную смену слоев кожи. Это единственный слой кожи, имеющий полноценную структуру и способный к делению). Дополнительная агрегация (процесс объединения элементов в одну систему) делящихся клеток аккумулирует прилегание к дистальному (наиболее удаленному от центральной точки) кончику отрубленного позвоночника, что позволяет впервые показаться бластеме (скопление неспециализированных клеток на раневой поверхности после ампутации органа). Продолжающийся рост бластемы происходит совместно с началом ангиогенеза (процесс образования новых кровеносных сосудов в органе или ткани), за чем следует новообразование периферических аксонов (длинный цилиндрический отросток нервной клетки), по которому нервные импульсы идут от тела клетки, и эпендимальной трубы позвоночника. Видоизменение костной ткани и новообразование мышц откладываются до того, как нарастание хвоста уже будет идти полным ходом.
Таким образом, потеря хвоста влечет за собой латентный период без-шрамного заживления раны, а регенерация регулируется бластемным делением. Таким образом, случаи значительной эпиморфной регенерации присущи в основном только позвоночным.
Известно также, что регенерация напрямую зависит от нервных окончаний. Центральная нервная система играет решающую роль в регенерации хвоста. Например, удаление спинного мозга рядом с местом отделения хвоста тормозит регенерацию, в то время как удаление наиболее близлежащих спинальных ганглий у корешков спинного мозга такого эффекта не имеет.
Данное исследование нацеливалось на преодоление эволюционной пропасти между млекопитающими, не склонными к успешной регенерации, и представителями не-амниотов, идеально овладевшими этой техникой. Процесс регенерации хвоста у ящериц – это высокоорганизованный феномен, заключающийся в полном, хотя и не идентичном восстановлении комплексной многотканевой структуры».
Анатомия хвоста: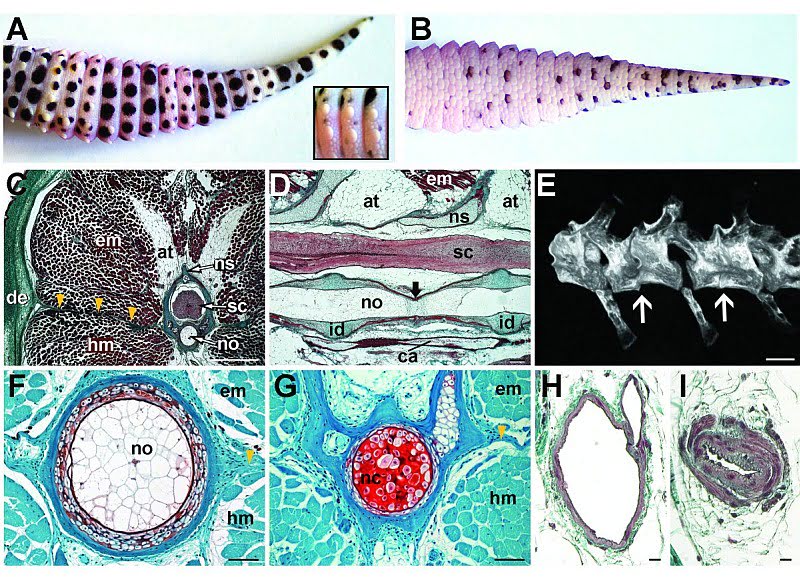
Размножение клеток в первоначальном хвосте: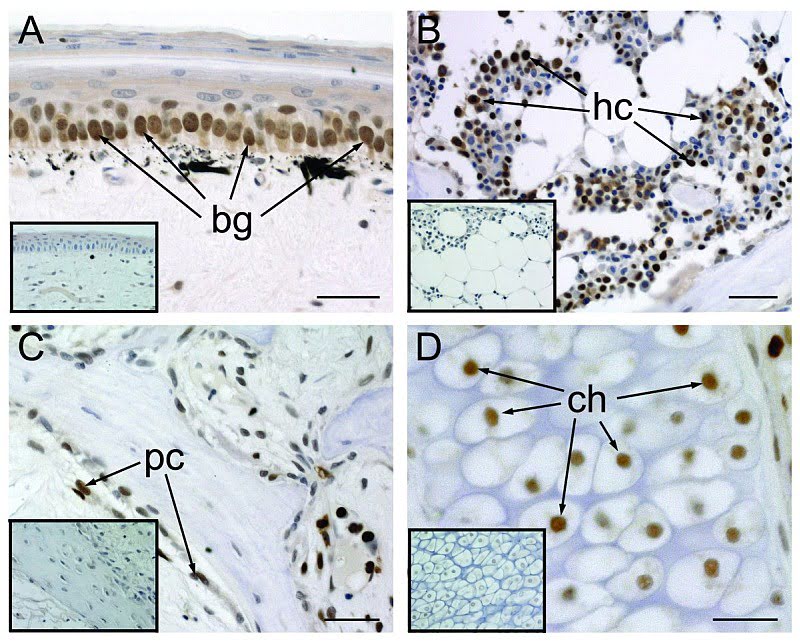
Регенерация хвоста:

Вторая стадия:
Третья стадия:

Четвертая стадия:

Пятая стадия:
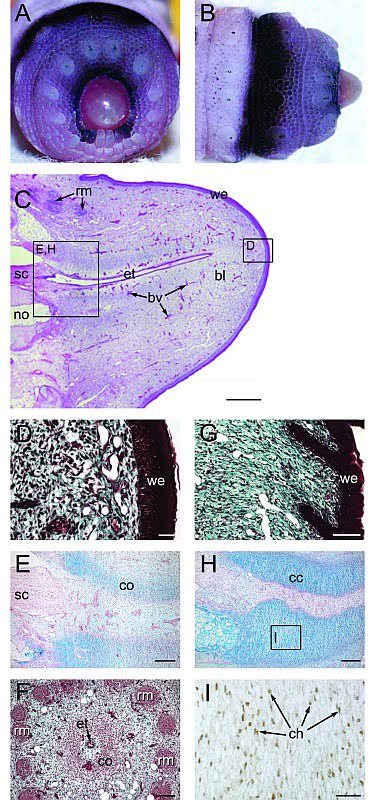
Шестая стадия:

Седьмая стадия:

Данная статья была составлена по материалам доклада «Новая амниотическая модель эпиморфной регенерации: леопардовый геккон» К. МакЛин и М. Викариуса и научной статьи из журнала экспериментальной биологии «Временные вариации управляемого движения отброшенных хвостов леопардовых гекконов: поведенческие модуляции» Т. Хигэла и Э. Рассела.
Перевод подготовлен для HGF Дарьей Орловой, 2016 год.